Nature Vol. 348 29 November 1990
Laboratory of Eukaryotic Molecular Genetics,
MRC National Institute for Medical Rewarch, The Ridgeway, Mill
Hill,
London NW7 1AA, UK
THE development of a eutherian mammal as a male is a consequence of testis formation in the embryo, which is thought to be initiated by a gene on the Y chromosome. In the absence of this gene, ovaries are formed and female characteristics develop1. Sex determination therefore hinges on the action of this testis-determining gene, known as Tdy in mice and TDF in humans2,3. In the past, several genes proposed as candidates for Tdy /TDF have subsequently been dismissed on the grounds of inappropriate location or expressions 4-9. We have recently described a candidate for Tdy, which maps to the minimum sex-determining region of the mouse Y chromosomel 10,11. To examine further the involvement of this gene, Sry, in testis development, we have studied its expression in detail. Fetal expression of Sry is limited to the period in which testes begin to form. This expression is confined to gonadal tissue and does not require the presence of germ cells. Our observations strongly support a primary role for Sry in mouse sex determination.
Male and female mouse embryos are morphologically indistinguishable until about 11.5 days post coitum (d.p.c.). The first visible sign of male development, the formation of testes from the 'indifferent' genital ridges, occurs within 24 hours, with the alignment of Sertoli cells into cords. Any candidate for Tdy should therefore be expressed in male genital ridge at or before this stage of development. Our initial observations suggested this to be the case for Sry (ref. 10). We wished to determine whether Sry transcription in the genital ridge is part of a broader temporal or spatial profile of expression suggesting a general role, or whether it correlates more specifically with testis differentiation. In particular, we wished to determine whether Sry expression in the genital ridge is due to somatic or to germ cells, as it is known that the latter are not required for testis development 12,13.
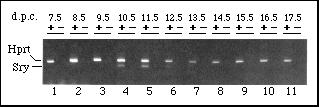
FIG. 1 Time course of fetal Sry expression.
RNA from mouse embryos of various stages was added to a 'reverse
transcription' reaction in the presence (+) or absence (-) of
reverse transcriptase (RT). Subsequent PCR reactions contained
oligonucleotide primers for hypoxanthine phosphoribosyltransferase
(Hprt ) and Sry. As expected, the 352-basepair (bp)-control
Hprt band was seen in all +RT samples. The 266-bp band
corresponding to Sry was only seen in 10.5 - 12.5-d.p.c.
samples. As the Sry primers are capable of amplifying genomic
DNA sequences, the absence of bands from all -RT samples confirms
that any signal is due to Sry transcripts and not to DNA
contamination.
METHODS. Small-scale RNA preparations were made, reverse transcribed
and amplified as previously described9,10 with the
addition of Perfect Match (Stratagene) to PCR reactions. Sry
primers 5'-3' were GAGAGC ATGGAG GGCCAT and CCACTC CTCTGT GACACT.
Annealing was at 53° C. Samples, divided into + and - RT fractions,
were from: 1, twelve whole embryos at 7.5 d.p.c.; 2, ten whole
embryos at 8.5 d.p.c.; 3, 0.1 µg of poly(A)+
RNA from pooled 9.5-d.p.c. embryos posterior to the forelimb-bud;
4, urogenital ridges from six 10.5-d.p.c. embryos; 5, urogenital
ridges from a single 11.5-d.p.c. embryo; 6, single 12.5-d.p.c.
testis; 7-11, 30, 25, 20,15 and 12% of single testes from 13.5-17.5-d.p.c.
fetuses, respectively. Parkes outbred mice were used unless otherwiw
stated.
RNA was extracted from mouse embryos and analysed for the presence of Sry transcripts by reverse transcription and polymerase chain reaction (PCR). No Sry expression was detected in 7.5-, 8.5- and 9.5-d.p.c. embryos, before genital ridge formation. Transcripts were first detected in the genital ridge at 10.5 d.p.c., were present at similar levels in the 11.5-d.p.c. urogenital ridge, and were less abundant in 12.5-d.p.c. testis (Fig. 1). Sry transcripts were not detected in testes from 13.5- to 17.5-d.p.c. Therefore, Sry expression corresponded precisely with the onset of testis differentiation, with a window of about one day either side.
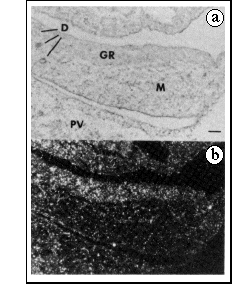
FIG. 2 In situ hybridization to a sagittal
section of an 11.5-d.p.c. male embryo, using an anti-sense RNA
probe for Sry.
a, Bright-field illumination. b, Dark-field illumination. GR,
genital ridge; M, mesonephros; D, mesonephric and paramesonephric
ducts; PV, prevertebrae. Scale bar, 50 µm.
METHODS. In situ hybridization was carried out as described22.
Embryos were fixed by staining for sex chromatin in amnion cells23.
[35S]UTP- labelled anti-sense RNA probe, corresponding
to the 374-bp Bgl II-Pst I fragment of p.4.2.2. (ref 10) was used.
This probe showed no specific labeling to sections of female embryos,
and control sense RNA probes did not show a specific signal on
male and female sections (data not shown). Autoradiographic exposure
was at 4° C for 6 days.
In situ hybridization was used to examine the distribution of Sr,v transcripts in 11.5-d.p.c. embryos. Although the level of expression was low, the hybridization signal was clearly associated with the genital rather than the adjacent mesonephric component of the urogenital ridge (Fig. 2). The homogeneity of the signal over the genital ridge indicates that the cells expressing Sry represent a large population, not confined to any specific region of the genital ridge, and not organized into specific structures. It was not possible to detect hybridization above background in 12.5-d.p.c. testes, after the formation of testis cords. The distribution of Sry transcripts in fetal tissues at 11.5-d.p.c. was further examined by PCR. When RNA was extracted from urogenital ridge, head, viscera and carcass fractions, there was no evidence for expression in any tissue other than urogenital ridge.
b
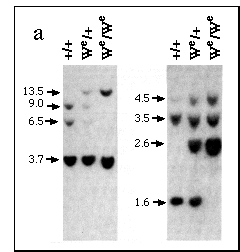
FIG. 3 Analysis of We embryos. a,
Restriction fragment length polymorphism (RFLP) seen on Southern
blots of DNA digested with Bgl II (left panel) or Xba I and Pst
I (right panel) using a probe for c-kit (refs 16,17); approximate
band sizes in kb are shown on the left.
b, PCR analysis of Sry expression in 11.5-d.p.c. urogenital
ridges of homozygous wild type (+) or mutant (W) embryos. Genotypes
were deduced by RFLP analysis. Left and right lanes for each sample
were +RT and -RT fractions, respectively (see Fig 1). Littermates
are bracketed as follows (1, 2, 3), (4, 5, 6), (7), (8, 9).
METHODS. a, DNA was digested, electrophoresed on 0.8% agarose-TBE
gels, and transferred to Hybond N filters (Amersham). DNA was
from wild type or heterozygote adults and from 14.5-d.p.c. fetuses
identified as homozygous mutant by liver morphology24.
Filters were hybridized with a probe corresponding to the 3' terminal
4.3-kb fragment of c-kit complementary DNA25,
labelled with 32P using a Multi-Prime kit (Amersham),
according to the manufacturer's instructions. After washing with
0.5X SSC buffer at 65° C, filters were autoradiographed for
16 h. Band sizes were estimated using l-Hind III markers.
b, Urogenital ridge RNA from individual progeny of C3H/H-101 We/+
matings was divided into +RT and -RT fractions and processed as
described in Fig. 1 legend.
In mouse fetuses homozygous for mutations at the dominant white spotting (W) locus, testes form normally despite a lack of germ cells12-14. If Sry is to be considered a candidate for Tdy, it ought to be expressed in We/ We as well as in wild-type genital ridges. We were able to identify embryos homozygous for the We allele15 by virtue of a restriction fragment length polymorphism detected using a probe for c-kit16,17 (Fig. 3a). In PCR analyses of Sry expression in genital ridge at 11.5 d.p.c., We/We fetuses proved indistinguishable from wild type (Fig. 3b).

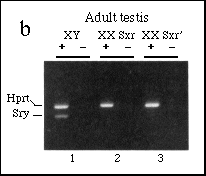
FIG. 4 Analysis of Sry expression in
fetal and adult tissues.
a, Northern blot analysis revealed a 1.3-kb transcript in adult
XY testis RNA (lane 4), but not in other adult tissues or in 11.5-d.p.c.
urogential ridge (lane 3), 12.5-d.p.c. male (or female) gonads
(lanes 1 and 2), or other adult tissues (lanes 5-10). Size (kb)
on the right.
b, PCR analysis of adult testis RNA. The 266-bp band corresponding
to Sry transcripts was seen in the normal XY testis sample
(1) but not XX Sxr or XX Sxr' samples (2, 3), whereas the control
Hprt band was seen in all samples. Reverse transcriptase
was included (+) or omitted (-) as described in Fig. 1.
METHODS. a, 20 µg of total RNA extracted from pools of fetal
or adult tissues was loaded in each lane, electrophoresed, transferred
to Gene-Screen (Dupont), and probed with an RNA probe labelled
with 32P to a specific activity of 1 x 109
c.p.m. µg-1. After hybridization, the filter
was treated with RNase to reduce non-specific background, then
exposed for 3 days.
b, Total RNA (0.2 µg) was divided into +RT and -RT fractions
and processed as described in Fig. 1 legend.
We were unable to detect the Sry transcripts present in 11.5d.p.c. genital ridge or 12.5-d.p.c. testis by northern blotting, presumably because of the low level of expression. Northern analysis of adult testis RNA revealed Sry transcripts of a similar size to those seen in adult human testis (Fig. 4a). In mouse testis, the transcripts are about 1.3 kilobases (kb) and run as a tight doublet under some conditions. To investigate whether the expression seen in adult testis was somatic, we used PCR to analyse RNA from XX Sxr and XX Sxr' adult mouse testes, which lack germ cells18,19. In contrast to the situation in fetal gonads, Sry expression in adult testis is dependent on germ cells (Fig. 4b). We are now addressing the question of whether this expression is a property of germ cells themselves, or a result of somatic-germ cell interaction. It is also unclear whether the adult and fetal transcripts are physically or functionally distinct. A number of genes suspected of having a role in directing developmental events in the embryo show reactivation in adult testis, but in no case has their function there been defined 20.
Sry has been shown to meet several criteria that can be applied to any candidate for Tdy. It maps to the appropriate area of the Y chromosome, is conserved in a wide range of mammals, is deleted in a line of XY female mice known to be mutant in Tdy, and encodes a product consistent with a regulatory role10,11,21. The present results show that Sry follows a tightly controlled spatial and temporal profile of expression that correlates well with testis differentiation. As this did not depend on the presence of germ cells, we conclude that Sry is expressed in one of the somatic cell lineages present in the developing gonad, as must be the case if Sry is to be considered a candidate for Tdy. The rapid cessation of Sry transcription after testis cord formation suggests that Sry initiates a cascade of gene expression, but is not required for the maintenance of gene activity in the developing testis. It will be interesting to determine the nature of the genes involved in subsequent steps of the pathway.
Received 25 September; Accepted 12 October, 1990.
REFERENCES
1. Jost, A., Vigier, B., Prepin, J. & Perrchellett, J. P. Recent Prog Horm Res. 29,1-41(1973).
2. McLaren, A. Trends Genet. 4, 153-157 (1988).
3. Goodfellow, P. N. & Darling, S. M. Development 102, 251-258 (1988).
4. Wachtel, S. S., Ohno, S.. Koo, G. C. & Boyse, E. A. Nature 257, 235-238 (1975).
5. MrLaren, A., Simpson, E., Tomonari, K., Chandier, P. & Hogg. H. Nature 312, 552-555 (1984).
6. Singh, L., Phillips, C. & Jones, K. W. Cell 36, 111-120 (1984).
7. Page, D. C. et al., Cell 51, 1091-1104 (1987).
8. Palmer, M. S. et al., Nature 342 937-939 (1989).
9 Koopman, P. et al., Nature 342, 940-942 (1989).
10. Gubbay, J. et al., Nature 346, 245-250 (1990).
11. Sinclair, A. H. et al., Nature 346, 240-244 (1990).
12. McLaren, A. in The Origin and Evolution of Sex (eds Halvorson H. O. & Monroy, A.) 289-300 (Liss, New York, 1985).
13. Merchant H. Devl Biol. 44, 1-21 (1975).
14. Mintz, B. & Russel, E. S. J. Exp. Zool. 134, 207-237 (1957).
15. Cattanach, B. M. Mouse News Lett. 59, 18 (1978).
16. Chabot, B., Stephenson, D. A., Chapman, V. M., Besmer, P. & Bernstein, A. Nature 335, 88-89 (1988).
17. Gessler, E. N., Ryan, M. A. & Housman, D. E. Cell 55, 185-192 (1988).
18. Cattanæh, B. M., Poljard, C. E. & Havskes, S. G. Cytogenetics 10, 318-337 (1971).
19. Burgoyne, P. S., Levy, E. R. & McLaren, A. Nature 320,170-172 (1988).
20. Woigemuth, D. J. et al., Proc. natn. Acad. Sci. U.S.A. 84, 5813-5817 (1987).
21. Lovell-Badge, R. H. & Robertson, E. Development 109, 635-646 (1990).
22. Wilkinson, D. G., Bailes, J. A. & McMahon, A. P. Cell 50, 79-88 (1987).
23. Monk, M. & McLaren, A. J. Embryol. exp. Morph. 63, 75-84 (1981).
24. Borghese, E. Acta Anat. 36, 185-220 (1959).
25. Qiu, F. et al., EMBO J. 7, 1003-1011 (1988).
ACKNOWLEDGEMENTS. We thank A. McLaren and M. Buehr for allowing us the use of We mouse stocks, P. Besmer for the c-kit probe, P. Goodfellow for critical comments, D. Wilkinson for assistance with in situ hybridization, and J. Gubbay. J. Collignon and A Economou for stimulating discussions. Also, the photography and oligonucleotide synthesis services at MRC-NIMR for their efficiency. A.M. is supported by the DAAD and Carl Duisberg Stiftung for das Auslandsstudium deutscher Studenten, and B.C. by the NIH.
Return To Molecular Biology Main Page
Return To Biology Dept. Main Page
![]()
©
Copyright 2000 Department of Biology, Davidson College, Davidson,
NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu