Cell, Vol. 61, 1359-1363, June 29, 1990. Copyright ©1990 by Cell Press
Michael J. Lewis, Deborah J. Sweet, and
Hugh R. B. Pelham
MRC Laboratory of Molecular Biology
Hills Road
Cambridge CB2 2QH
England
Summary
Luminal ER proteins carry a signal at their C terminus that prevents their secretion; in S. cerevisiae this signal is the tetrapeptide HDEL. Indirect evidence suggests that HDEL is recognized by a receptor that retrieves ER proteins trom the secretory pathway and returns them to the ER, and a candidate for this receptor is the product of the ERD2 gene (see accompanying paper). We show here that presumptive ER proteins from the budding yeast K. lactis can terminate either with HDEL or, in the case of BiP, with DDEL. S. cerevisiae does not efficiently recognize DDEL as a retention signal, but exchange of its ERD2 gene for the corresponding gene from K. lactis allows equal recognition of DDEL and HDEL. Thus the specificity of the retention system is determined by the ERD2 gene. We conclude that ERD2 encodes the receptor that sorts luminal ER proteins.
Introduction
Proteins that reside in the lumen of the ER are characterized
by a C-terminal signal that prevents their passage along the secretory
pathway (for review see Pelham, 1989). The optimal signal in animal
cells is the tetrapeptide KDEL, whereas in Saccharomyces cerevisiae
it is HDEL. Proteins bearing this retention signal are able to
leave the ER by vesicular transport but are specifically retrieved
from the Golgi or a pre-Goigi "salvage" compartment.
In yeast, the retrieval system is readily saturable; it is presumed
to involve a membrane-bound receptor that cycles between the Golgi
and the ER (Dean and Pelham, 1990).
In an attempt to identify components of this sorting system, and in particular the presumed HDEL receptor, we have isolated yeast mutants that fail to retain HDEL-tagged proteins in the ER (Pelham et al., 1988; Hardwick et al., 1990). These mutants define two genes, one of which (ERD2)is a good candidate for the receptor. In support of this, we have found that the capacity of the retention system is controlled by the level of ERD2 expression: low levels of ERD2 protein result in secretion of ER proteins, while high levels render the system resistant to saturation (Semenza et al., 1990).
The definitive property of a receptor is its ligand specificity. Thus one way to test whether ERD2 encodes the receptor is to identity differences in the specificity of the retention system between species, and see whether they are determined by the ERD2 gene. We report here that one of the major ER proteins of the budding yeast Kluyveromyces lactis , namely BiP, has at its C terminus the sequence DDEL, although at least one glycoprotein in this species carries the HDEL signal. S. cerevisiae does not efficiently retain fusion proteins that terminate with DDEL, but replacement of the ERD2 gene with the equivalent one from K. lactis allows efficient retention of both DDEL and HDEL. This result argues very strongly that the ERD2 protein is indeed the receptor responsible for retrieving ER proteins from the Golgi.
Results
K. lactis BiP Lacks HDEL
We have raised anti-peptide antisera that are specific for a C-terminal
HDEL sequence. These antibodies recognize two different peptides
whose only common sequence is HDEL (Hardwick et al., 1990), but
will not bind to HDEL when it is present at an internal position,
nor to KDEL. In a search for a suitable organism whose ER proteins
have a signal different from those in S. cerevisiae, we
used these antibodies to probe blots of total proteins from the
yeasts Schizosaccharomyces pombe and Kluyveromyces lactis
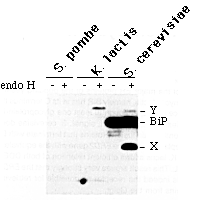
(Figure 1). K. lactis had at least one glycoprotein that
was detected by the antiserum; from its mobility and abundance,
it seems likely that this glycoprotein corresponds to the grp94
protein of animal cells. A smaller, unglycosylated protein was
also detected (Figure 1). However, only an extremely faint signal
was detected in the position expected for K. lactis BiP,
implying that it carries a divergent retention signal. S. pombe
contained no abundant proteins that reacted with the antiserum
(Figure 1), and thus this species must also use a retention signal
other than HDEL.
Figure 1. Divergence of the ER Retention Signal
in Different Yeasts
Samples of total protein from the indicated yeasts were incubated
with endoglycosidase H to remove high mannose oligosaccharides.
Endoglycosidase H-treated and control samples were immunoblotted
with anti-HDEL antibodies. X and Y refer to the glycoproteins
previously identified in S. cerevisiae; Y probably corresponds
to grp94 (Hardwick et al., 1990). Staining of the blot revealed
similar amounts of protein in each lane.
K. lactis and S. cerevisiae are related budding yeasts, and genes from one are usually efficiently transcribed and correctly spliced in the other (e.g., Salmeron and Johnston, 1986; Desher et al., 1989); in contrast, many S. pombe genes are nonfunctional in S. cerevisiae. K. lactis thus seemed a suitable organism with which to test the hypothesis that the ERD2 gene encodes the HDEL receptor.
Cloning the K. lactis BiP Gene
To identify the retention signal on K. lactis BiP, we cloned
the corresponding gene. Low stringency hybridization with parts
of the S. cerevisiae BiP gene failed to distinguish the
K. lactis BiP gene from other members of the hsp70 family.
K. lactis DNA was therefore digested with EcoRI and electrophoresed
on an agarose gel, and DNA from regions corresponding to individual
cross-hybridizing bands was eluted, cloned, and sequenced. The
third fragment sequenced showed clear homology with BiP and was
used to isolate a full-length clone. From this, we obtained sequence
corresponding to the C terminus of the protein (Figure 2).
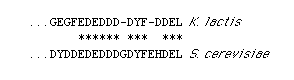
Figure 2. C-Terminal Sequences of K. lactis
and S. cerevisiae BiP Proteins
Asterisks indicate identical residues; two gaps have been introduced
into the K. lactis sequence to maximize homology The complete
sequence of the K. lactis gene will be reported elsewhere.
Strikingly, the C-terminal tetrapeptide of K. lactis BiP is DDEL. The presence of an aspartic acid in place of the histidine found in S. cerevisiae accounts for the failure of our anti-HDEL antibodies to recognize the protein.
Cloning of K. lactis ERD2
Probing Southern blots of K. lactis DNA at low stringency
with the S. cerevisiae ERD2 gene revealed a single
cross-hybridizing sequence, which could be localized to a 1.25
kb EcoRI-HindIII fragment. This fragment was cloned and sequenced
(Figure 3A). The gene contained a single intron in the same location
as the intron in the S. cerevisiae gene, and the encoded
proteins showed 59% identity, with many of the changes being conservative
in nature. Furthermore, expression of the K. lactis gene
in S. cerevisiae was sufficient to complement the growth-arrest
phenotype of an ERD2 deletion mutant. It thus seems clear
that the K. lactis gene that we have isolated corresponds
to the ERD2 gene of S. cerevisiae.

B

Figure 3. Sequence of K. lactis ERD2
(A) Nucleotide sequence of the gene. Underlining indicates important
features: the initiation codon; splice donor, branchpoint, and
acceptor; and termination codon. The sequence illustrated is sufficient
for full ERD2 function.
(B) Alignment of the S. cerevisiae (upper) and K. lactis
(lower) ERD2 protein sequences. Asterisks indicate identical
residues, and charged amino acids are indicated. Two spaces have
been introduced into each sequence to maximize homology. Bars
above the S. cerevisiae sequence indicate a 15 amino acid
duplication.
A comparison of the two sequences is shown in Figure 3B. One difference is that the 15 amino acid duplication near the N terminus of S. cerevisiae ERD2is barely recognizable as a repeated sequence in K. lactis. Moreover, the D --> N change at amino acid 51, which causes a strong erd phenotype in S. cerevisiae, is naturally present in the heterologous gene. At the C terminus, three hydrophobic stretches are very highly conserved, although the hydrophilic sequences flanking them are more divergent. These hydrophobic residues may form part of a conserved transmembrane structure.
Changing the Specificity of the Retention System
To determine the effects of ERD2 on the specificity of
the retention system, we expressed fusion proteins carrying at
their C termini the potential retention sequences FEHDEL (corresponding
to S. cerevisiae BiP), YFDDEL (K. lactis BiP), or
SEKDEL (rat BiP). Control constructs lacking these sequences terminated
with SEEDLN. Initial experiments used a fusion protein containing
the enzyme invertase. Secretion of this protein can easily be
monitored by assaying unlysed cells for invertase activity; intracellular
enzyme is inaccessible to substrate under these conditions, but
can be detected after cell lysis.
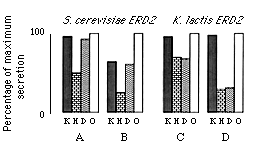
Figure 4. Effect of ERD2 on the Secretion
of Invertase Fusion Proteins Bearing Different C Termini
The percentage of total invertase activity that had been secreted
was determined for fusion proteins terminating with SEEDLN (O),
or with the same sequence followed by SEKDEL (K), FEHDEL (H),
or YFDDEL (D).
(A) Wild-type S. cerevisiae strain with its normal chromosomal
copy of ERD2.
(B-D) Isogenic strains that have the chromosomal ERD2 gene
deleted, and carry the following plasmids: a high copy number
plasmid (PER220) with the S. cerevisiae ERD2 gene
(B), a low copy number plasmid (LE22) expressing K. lactis
ERD2 (C), and a high copy number plasmid (LE21) with the
K. lactis ERD2 gene (D).
Values (average of four or more determinations) have been normalized
to the control construct (O) to aid comparison; this corrects
for minor differences in overall secretion efficiency, which reflect
the slightly different growth rates of the strains. About 30%
of the invertase activity was secreted from the control strains.
As shown in Figure 4A, the HDEL- tagged fusion protein was secreted less efficiently from a wild-type strain than were the KDEL or DDEL versions, neither of which showed significant retention relative to the control. Thus, neither KDEL nor DDEL is efficiently recognized by the S. cerevisiae receptor under these conditions.
A slightly different result was obtained with a strain that lacked the chromosomal copy of ERD2 but instead carried the gene on a multicopy vector. Retention of the HDEL construct was improved, but in addition partial retention of both DDEL and KDEL was observed (Figure 4B). Thus when the capacity of the system is increased by overexpression of ERD2, its apparent specificity is broadened. Presumably, under normal conditions the receptor is saturated with endogenous HDEL-containing proteins, which effectively compete with the DDEL and KDEL sequences, but when receptor is present in excess, a weak affinity for the latter sequences is revealed. Nevertheless, with both high and low levels of ERD2 expression, HDEL was the preferred substrate.
A different result was obtained when the plasmid-borne ERD2 gene was replaced with K. lactis ERD2. Two different plasmids expressing the K. lactis gene were used; one was a low copy number vector with the ERD2 coding sequences fused to the TPI promoter, while the other was a high copy number plasmid carrying the gene with its own promoter. Strains containing these plasmids showed different efficiencies of retention, presumably because they expressed ERD2 at different levels, but in each case the specificity was the same: invertase fusion proteins bearing HDEL and DDEL were retained equally well, while the KDEL construct was secreted as efficiently as the control (Figures 4C and 4D).
As a further test, we examined the secretion of prepro-a factor fusion proteins in strains lacking
the chromosomal ERD2 gene and carrying either S. cerevisiae
or K. lactis ERD2 on a multicopy plasmid. Pro-a factor is secreted rapidly, and the fusion
proteins can most easily be assayed by pulse-labeling and immunoprecipitation.
Molecules that pass through the Golgi are proteolytically processed
and thus escape detection, so the amount of labeled protein remaining
after a short chase gives a direct indication of the efficiency
of retention (Dean and Pelham, 1990; Semenza et al., 1990).
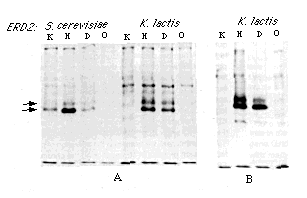
Figure 5. Effect of ERD2 on Secretion
of Pro-a
factor Fusion Proteins
Fusion proteins bearing the same termini as those in Figure 4
were assayed by pulse-labeling and immunoprecipitation.
(A) Strains carrying the chromosomal ERD2 deletion and
high copy number plasmids expressing either S. cerevisiae
or K. lactis ERD2 were labeled for 10 min and chased
for 15 min.
(B) Deletion strains carrying the low copy number plasmid expressing
the K. lactis ERD2 gene were labeled for 10 min
and chased for 5 min.
Arrows indicate pro-a factor The lower band is the ER form, and the upper band corresponds to protein that has cycled through an early Golgi compartment.
We conclude from these experiments that the ERD2 gene controls the specificity of the ER retention system. S. cerevisiae ERD2 allows strong recognition of HDEL and weak recognition of DDEL and KDEL; K. lactis ERD2 allows recognition of HDEL and DDEL, but not KDEL. The dual specificity of K. lactis ERD2is consistent with the presence of both DDEL and HDEL on ER proteins of this organism.
Discussion
Our aim in these experiments was to test the hypothesis that the
ERD2 gene encodes the receptor that sorts luminal ER proteins.
We first looked for a species of yeast with a receptor specificity
different from that in S. cerevisiae, then isolated its
ERD2 gene, transferred it to S. cerevisiae, and
tested for a change in the specificity of the retention system.
The results indicate that the pattern of retention is indeed determined
by the ERD2 gene.
Divergence of the Retention Signal for Luminal ER Proteins
We were surprised to find that the BiP protein of K. lactis
had the C-terminal sequence DDEL, because it implies considerable
divergence of the retention signal between two related budding
yeasts. All known examples of luminal ER proteins in S. cerevisiae
have HDEL at their C terminus; these include BiP (Rose et al.,
1989; Normington et al., 1989; Nicholson et al., 1990), protein
disulfide isomerase (M. F. Tuite and R. Freedman, personal communication),
the KRE5 gene product (Meaden et al., 1990), and two other proteins
identified only by their reactivity with anti-HDEL antibodies.
Proteins from other species, including plants, nematodes, insects,
and vertebrates, almost invariably have a positively charged amino
acid 4 residues from the terminus (reviewed by Pelham, 1989),
although Plasmodium falciparum BiP has serine (Kumar et
al., 1988). The occurrence of an acidic residue at this position
in K. lactis BiP is thus unprecedented.
Despite this striking difference, at least one glycoprotein in K. lactis has an HDEL sequence, as judged from its ability to bind anti-HDEL antibodies. The presence of two rather different sequences raises the question of how recognition of both of them is achieved.
ERD2 Encodes the Sorting Receptor
Previous experiments had identified the ERD2 protein as
a good candidate for the HDEL receptor, because its abundance
controls the capacity of the retention system (Semenza et al.,
1990). We have shown here that the signal specificity of the system
is also determined by ERD2. With S. cerevisiae ERD2,
HDEL is the preferred signal, whereas when the gene is replaced
by the K. lactis homolog, HDEL and DDEL are retained with
equal efficiency. This difference is independent of the level
of expression of the ERD2 genes and thus cannot be explained
by a mere quantitative difference in the activity of the two ERD2
proteins. Furthermore, the observed preferences are entirely consistent
with the known sequences of ER proteins in the two species. We
therefore conclude that ERD2 indeed encodes the receptor.
In the case of K. lactis, this single receptor must be capable of binding both FEHDEL and YFDDEL, which suggests a surprising degree of flexibility in its interactions. The specificity of the S. cerevisiae receptor is also not absolute, because when it is expressed at high levels there is some recognition of both DDEL and KDEL sequences, neither of which has been found in this species. Presumably, this reflects weak binding of the receptor to DEL. The dual specificity of the K. lactis receptor cannot, however, be explained simply by binding to DEL, because SEKDEL is not recognized.
One possibility that we cannot rule out is that binding of YFDDEL to the K. lactis receptor is influenced by the two aromatic residues upstream of the terminal tetrapeptide, but there is no evidence that more than 4 amino acids are recognized in other species. We therefore prefer a model in which HDEL and DDEL each interact with the receptor in a highly specific manner.
The ERD2 protein not only functions as a receptor, but is also required for normal vesicular traffic through the Golgi (Semenza et al., 1990). If ligand binding involves parts of the protein that are also required for viability, it will be difficult to identify the binding site by mutation. The results in this paper suggest that it may be possible to identify the sequences responsible for ligand specificity by creating chimeric ERD2 genes with sequences from S. cerevisiae and K. lactis. It should also be possible to identify conserved functional motifs by isolating the receptor gene from more distantly related species. Very recently, an anti-idiotypic antibody approach has identified a 72 kd glycoprotein that is good candidate for the KDEL receptor in animal cells (Vaux et al., 1990). It will be interesting to see whether part of this protein shows similarity to the 26 kd ERD2 product.
Experimental Procedures
Strains
Derivatives of S. cerevisiae strain SEY2102 (MATa suc2-d9
ura3-52 /eu2-3,-112 his4-519) were used for Figure 4A. Other
results were obtained with derivatives of the ERD2 deletion
strain D209 (Semenza et al., 1990). K. lactis was kindly
provided by Stephen Johnston (Duke University), and S. pombe by
Andrew Newman.
Cloning
The K. lactis BiP and ERD2 genes were detected by
hybridization with probes containing the corresponding S. cerevisiae
coding sequences, which we had isolated previously (Hardwick et
al., 1990; Semenza et al., 1990). Hybridization was in 4x SSC,
10% formamide at 42°C, and filters were washed with 2x SSC
at 50°C. Fragments migrating with the hybridizing bands were
excised from an agarose gel and cloned into M13 vectors. The relevant
clones were identified in these minilibraries by plaque hybridization.
For high level expression, the EcoRI-HindIII fragment containing the K. lactis ERD2 gene was inserted into the LEU2-containing 2µm vector ZUC13 to form plasmid LE21. Expression at a lower level was obtained by fusing the gene at the Scal site (position 280) to the TPI promoter and inserting it into a vector containing TRP1, ARS1, and CEN3 to form plasmid LE22.
Isogenic strains carrying a deletion of the chromosomal ERD2 gene and the K. lactis ERD2 plasmids LE21 or LE22, or the S. cerevisiae ERD2 plasmid PER220, were generated as described by Semenza et al. (1990).
Invertase and pro-a factor fusion constructs were identical to those used previously (Pelham et al., 1988; Dean and Pelham, 1990) except that additional constructs encoding proteins terminating with YFDDEL were prepared using synthetic oligonucleotides. All were inserted at the URA3 locus. For the experiments shown in Figure 5, only strains containing a single inserted gene (identified by Southern blot analysis) were used. Invertase assays, pulse labeling, and immunoprecipitation are described elsewhere (Pelham et al., 1988; Dean and Pelham, 1990). Note that the 2 factor constructs contain the epitope recognized by the monoclonal antibody 9E10, allowing their specific immunoprecipitation without interference from endogenous 2 factor
Acknowledgments
We thank Kevin Hardwick for performing the experiment shown in Figure 1, Jan Semenza, Stephen Johnston, and Andrew Newman for yeast strains, Neta Dean for the pro-a factor plasmids, Mark Bretscher, Barbara Pearse, and Sean Munro for helpful comments on the manuscript, and Steve Fuller, David Vaux, and John Tooze for communication of their results prior to publication.
The costs of publication of this article were
defrayed in part by the payment of page charges. This article
must therefore be hereby marked "advertisement"
in accordance with 18 U.S.C. Section 1734 solely to indicate this
fact.
Received May 10, 1990.
References
Dean, N., and Pelham, H. R. B. (1990). Recycling of proteins from the Golgi compartment to the ER in yeast. J. Cell Biol., in press.
Desher, J. O., Larson, G. P, and Rossi, J. J. (1989). Kluyveromyces lactis maintains Saccharomyces cerevisiae intron-encoded splicing signals. Mol. Cell. Biol. 9, 2208-2213.
Hardwick, K. G., Lewis, M. J., Semenza, J., Dean, N., and Pelham, H. R. B. (1990). ERD1, a yeast gene required for the retention of luminal endoplasmic reticulum proteins, affects glycoprotein processing in the Golgi apparatus. EMBO J. 9, 623 630.
Kumar, N., Syin, C., Carter, R., Quakyi, I., and Miller, L. H. (1988). Plasmodium falciparum gene encoding a protein similar to the 78-kDa rat glucose-regulated stress protein. Proc. Natl. Acad. Sci. USA 85, 6277-6281.
Meaden, R, Hill, K., Wagner, J., Slipitez, D., Sommer, S. S., and Bussey, H. (1990). The yeast KRE5 gene encodes a probable endoplasmic reticulum protein required for (1-6)-ß-D-glucan synthesis and normal cell growth. Mol. Cell. Biol. 10, 3013 3019.
Nicholson, R. C., Williams, D. B., and Moran, L. A. (1990). An essential member of the hsp70 gene family of Saccharomyces cerevisiae is homologous to immunoglobulin heavy chain binding protein. Proc. Natl. Acad. Sci. USA 86, 1159 1163.
Normington, K., Kohno, K., Kozutsumi, Y, Gething, M.-J., and Sambrook, J. (1989). S. cerevisiae encodes an essential protein homologous in sequence and function to mammalian BiR Cell 57, 12231236.
Pelham, H. R. B. (1989). Control of protein exit from the endoplasmic reticulum. Annu. Rev. Cell Biol. 5, 1-23.
Pelham, H. R. B., Hardwick, K. G., and Lewis, M. J. (1988). Sorting of soluble ER proteins in yeast. EMBO J. 7; 1757-1762.
Rose, M. D., Misra, L. M., and Vogel, J. R (1989). KAR2, a karyogamy gene, is the yeast homolog of the mammalian BiP/GRP78 gene. Cell 57, 1211-1221.
Salmeron, J. M., and Johnston, S. A. (1986). Analysis of the Kluyveromyces lactis positive regulatory gene LAC9 reveals functional homology to, but sequence divergence from, the Saccharomyces cerevisiae GAL4 gene. Nucl. Acids Res. 14, 7767-7781.
Semenza, J. C., Hardwick, K. G., Dean, N., and Pelham, H. R. B. (1990). ERD2, a yeast gene required for the receptor-mediated retrieval of luminal ER proteins from the secretory pathway. Cell 61, this issue.
Vaux, D., Tooze, J., and Fuller, S. (1990). Network antibodies identify an intracellular membrane protein that recognises the mammalian ER retention signal. Nature 345, 495-502.
GenBank Accession Number
The accession number for the sequence reported in this paper is M34844.
![]()
![]()
©
Copyright 2000 Department of Biology, Davidson College, Davidson,
NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu