This is a web page for an undergraduate course at Davidson College.
Term Paper on Rheumatoid Arthritis
Histamine
Elizabeth Super
Dr. Malcolm Campbell
Biology 307: Immunology
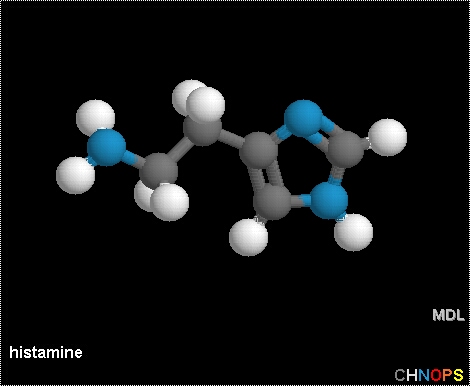
Fig. 1 Structural representation of histamine. Histamine, or 2-(4-imidazolyl)-ethyl-amine, is a dibasic vasoactive amine that is located in most body tissues but is highly concentrated in the lungs, skin, and gastrointestinal tract. A single polypeptide chain protein, histamine is stored in mast cells and basophils (Metcalfe et al., 1997).
Permission to use this image requested from vlesko@ccbc.cc.md.us on 18 March 2003. http://student.ccbc.cc.md.us/~cminnier/sample/Lectures/16LECT.htm.
Overview of Histamine (Rang et al., 1995)
1. Histamine is a basic amine that is stored in mast cells and basophils.
2. Histamine is held by ionic forces within intracellular granules by macroheparin.
3. The interaction between allergen and IgE, bound to the surface of mast cells
or
basophils by a surface receptor that binds the Fc fragment of IgE, leads to
degranulation of these cells, with release of mediators, including histamine.
4. It acts on H1-, H2-, or H3-receptors on target cells to produce its effects.
5. Its main actions include stimulation of gastric secretion, contraction of
most smooth
muscle (other than blood vessel smooth muscle), cardiac stimulation, vasodilation,
and increased vascular permeability.
6. When injected intradermally, histamine causes the “triple response”,
which includes
local vasodilation, wheal, and flare.
7. Histamine has two major pathophysiological roles, which include the stimulation
of
gastric acid secretion, which is treated with H2-receptor antagonists, and the
mediation of type 1 hypersensitivity reactions such as urticaria and hay fever,
which is treated with H1-receptor antagonists.
Overview of Mast Cells and Basophils
Mast cells and basophils are the effector cells involved in the immediate hypersensitivity
response (Bach, 1982). Found in tissues throughout the body, they are particularly
associated with blood vessels and nerves and are in proximity to surfaces that
border the external environment (Metcalfe et al, 1997). Both contain numerous
osmophilic granules that contain heparin and other proteins that support mediators,
including histamine, which alters cellular and vascular reactions. Secretion
of mediators occurs by degranulation during which the contents of the granules
are exocytosed. Degranulation is provoked by certain chemical agents, C3a and
C5a (two complement components) binding to surface receptors, certain drugs,
and the IgE system (Bach, 1982). Mast cells and basophils have receptors for
IgE antibodies and can be activated to secrete mediators if IgE first binds
to these receptors, followed by antigen binding to the Fab fragment of the fixed
IgE molecules (Rang et al., 1995). Degranulation can lead to allergic reactions
or anaphylactic shock, in extreme cases (Sompayrac, 1999).
Synthesis and Storage of Histamine
Histamine, or 2-(4-imidazolyl)-ethyl-amine, is a dibasic vasoactive amine that
is located in most body tissues but is highly concentrated in the lungs, skin,
and gastrointestinal tract. It is produced by the decarboxylation of histidine
by histidine decarboxylase (Paul, 1984). This reaction takes place in the Golgi
apparatus of mast cells and basophils (Metcalfe et al., 1997). A single polypeptide
chain protein, histamine is then stored in mast cells and basophils, where it
is localized primarily in the cytosol. In both mast cells and basophils, histamine
is detained in intracellular granules complexed with an acidic protein and macroheparin
that interact with the basic histamine by ionic forces (Rang et al., 1995).
Approximately 3-8 pg histamine/cell is found in mast cells that are isolated
from human lung, skin, lymphoid tissue, and the small intestine. Histamine dissociates
from its complex by cation exchange with extracellular sodium at a neutral pH
(Metcalfe et al., 1997). Therefore, it is readily available upon cell activation.
The granules comprise up to 70% of the weight of a mast cell, so substantial
extracellular histamine concentrations can be produced locally upon histamine
release. Histamine is less potent than other mediators linked to acute hypersensitivity,
so high local concentrations are required to produce significant effects in
vivo (Paul, 1984).
Release of Histamine
Mast cells release histamine during inflammatory or allergic reactions. When
exposed to allergens, humans produce IgE antibodies that are directed against
the allergen. Mast cells have receptors that can bind to the Fc region of IgE
antibodies (Sompayrac, 1999). IgE binds with high affinity and high specificity.
No conformational changes of the receptor occur when IgE binds. Binding of monomeric
IgE to the Fc receptor does not cause mast cell activation and degranulation;
initial exposure to an allergen causes large numbers of IgE antibodies to attach
to mast cells surfaces without activating them (Metcalfe et al., 1997). Upon
second exposure to an allergen, the IgE molecules bound to the mast cell surface
can bind to allergen. The allergen causes crosslinking of the IgE molecules
on the cell surface, which clusters the Fc receptors. This clustering stimulates
a signal transduction that causes the mast cell to ‘degranulate’,
or dump their granules into the tissues (Sompayrac, 1999).
Ishizaka and Ishizaka established that at least two specific IgE molecules, complexed with allergen, were required to induce skin reactions in man. Therefore, the important step in mast cell activation appears to be the crosslinking of IgE molecules, which causes the clustering of Fc receptors. This step also appears necessary because the bridging of receptors forms hydrophilic channels that allow calcium levels to increase intracellularly, which triggers mediator release. It has also been established that IgE acts as an anchor for antigen and helps to amplify the transduction signal generated by the bridging due to antigen-binding (Froese, 1980).
Histamine’s small size, low molecular weight, and diffusability cause
it to disperse from local sites, which limits its effect to a small period of
time (Paul, 1984). Specific receptors on target cells mediate the biological
activities of histamine (Metcalfe et al., 1997). Extracellular histamine is
rapidly metabolized, by enzymes present in leukocytes, to metabolites that are
excreted in the urine (Paul, 1984).
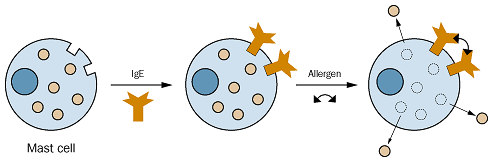
Fig. 2 Degranulation of a Mast Cell. Mast cells detain histamine in intracellular granules. Binding of IgE to cell surface receptors on a mast cell primes the cell to respond to allergen. Introduction of allergen and its subsequent binding to IgE induces crosslinking of IgE and clustering of Fc receptors. Clustering initiates a signal transduction event that stimulates the mast cell to degranulate, or release the contents of its granules. Mediators, such as histamine, which is represented by the tan circle, are released from granules and can bind to specific receptors to carry out their actions (Sompayrac, 1999).
Permission to use this image requested from feedback@chemsoc.org on 18 March 2003. http://www.chemsoc.org/chembytes/ezine/2000/eic_mar00.htm.
Histamine Receptors
Histamine acts by binding to receptors on target cells, and different cells
types express different receptor types. Upon binding to target cell receptors,
histamine causes intracellular events, such as phosphatidylinositol breakdown
to IP3 and DAG, which lead to different effects in different cell types (Abbas
et al., 1994). Three main types of histamine receptors – H1, H2, and H3
– have been identified and were distinguished by the actions of agonist
and antagonist drugs on each receptor type. Pyrilamine, cimetidine, and thioperamide
are selective antagonists at H1-, H2-, and H3-receptors, respectively. Dimaprit
and (R)a-methyl histamine are selective agonists for H2- and H3-receptors, respectively.
No selective agonists have been identified for H1-receptors. Histamine H1 antagonists
(commonly called antihistamines) and H2 antagonists have clinical uses, but
those acting at H3 receptors are currently utilized as research tools (Rang
et al., 1995). Antihistamines inhibit the wheal and flare response caused by
intradermal allergens or anti-IgE antibodies (Abbas et al., 1994).
H1-receptors are located in human bronchial muscle and are linked to transduction
systems that cause increased intracellular Ca2+, which leads to muscle contraction.
H2-receptors are located in acid-secreting stomach cells and in the heart. Stimulation
leads to gastric acid secretion and increased atrial rate. This receptor type
is linked to transduction systems involved in activation of adenylate cyclase
and increased production of cyclic AMP. H3-receptors are related to neural tissue
and are found at presynaptic sites; stimulation causes the inhibition of neurotransmitter
release (Rang et al., 1995).
Actions
Histamine produces many of the effects of inflammation and hypersensitivity,
including vasodilation, edema, increased vascular permeability, and smooth muscle
contraction (Rang et al., 1995). Increased vascular permeability causes fluid
to escape from capillaries into the tissues, which leads to the classic symptoms
of an allergic reaction – a runny nose and watery eyes (Sompayrac, 1999).
It is thought to be a major mediator of the acute inflammatory response, although
histamine H1 antagonists have little effect on acute inflammation (Rang et al.,
1995).
Gastric Secretion: The most important action of histamine, in a clinical sense, is its stimulation of gastric acid secretion by acting on H2-receptors. It is implicated in the formation of peptic ulcers (Rang et al., 1995).
Smooth Muscle Effects: Histamine causes contraction of the smooth muscle of the ileum, bronchi and bronchioles, and uterus by acting on H1-receptors. It may be involved in increased peristalsis associated with food allergies (Abbas et al., 1994). Histamine-induced bronchiolar constriction has been implicated in the first phase of bronchial asthma (Rang et al., 1995). In asthmatics, histamine was found to increase airway smooth muscle tone and cause mucosal edema and glandular secretion, resulting in the narrowing of the airways and limited air flow. In nonasthmatics, bronchial activity to histamine was limited, most likely due to fewer H1-receptors in airway smooth muscle (Goldie, 1990).
Cardiovascular Effects: Acting on H1-receptors, histamine causes the dilation of blood vessels; it induces endothelial cells to synthesize vascular smooth muscle relaxants, including prostacyclin and nitric oxide, which cause vasodilation (Abbas et al., 1994). Acting on H2-receptors, it increases heart rate and cardiac output. When injected intradermally, histamine leads to reddening of the skin, wheal, and flare, called the “triple response”. Vasodilation of small arterioles and precapillary sphincters causes reddening, while increased permeability of postcapillary venules causes the wheal; both these effects are implicated in activation of H1-receptors. Histamine does not increase capillary permeability. Histamine also induces an “axon reflex”, which leads to stimulation of sensory nerve fibers and the release of a vasodilator mediator; this causes the flare (Rang et al., 1995).
Itching: If histamine is injected into the skin, it causes itching, due to stimulation of sensory nerve endings (Rang et al., 1995).
Effects on Nasal Mucosa: Allergens can bind to IgE-loaded mast cells in the nasal mucosa, which leads to three clinical responses: sneezing results from histamine-associated sensory neural stimulation; hypersecretion from glandular tissue occurs; nasal mucosal congestion results due to vascular engorgement associated with vasodilation and increased capillary permeability (Monroe et al., 1997).
Role of Histamine in the Immune Response
Histamine was originally considered to be a mediator involved in the immediate
hypersensitivity response. It has also been shown to affect leukocyte function
and migration. It is involved in inhibition of lectin- or antigen-induced proliferation
of T cells, release of lymphokines from T cells, the induction of cytotoxic
T cells, cytolysis by mature cytotoxic T cells, B cell differentiation, lysosomal
enzyme release in neutophils, IgE-mediated histamine release from basophils,
and in chemokinetic effects on neutophils and eosinophils (Gallin et al., 1980).
These actions can be blocked by H2 antagonists. Many of these effects on leukocyte
function are inhibitory and can be seen as anti-inflammatory actions, which
can limit antibody hypersensitivity. However, H1 effects of histamine on blood
vessels and skin are proinflammatory and occur during hypersensitivity reactions.
It has been proposed that histamine initially promotes and later inhibits immune
responses (Paul, 1984).
Antihistamines
The study of histamine-receptor antagonists began nearly seventy years ago.
In 1937, Bovet and Staub discovered the first H1-receptor antagonist, which
marked the first generation of antihistamines to treat allergic diseases. Although
widespread in use, major CNS adverse effects, such as sedation and performance
deficits, and their anticholinergic activities caused doubt about their effectiveness.
A second generation of antihistamines was quickly developed that showed fewer
sedating side effects. The number of second-generation antihistamines has grown
rapidly, and their efficacies can be compared by injecting histamine epicutaneously
and viewing which antihistamines block the wheal (swelling) and flare (vasodilation)
response. Antihistamines suppress the histamine-induced wheal and flare response
by blocking the binding of histamine to its receptors on nerves, vascular smooth
muscle, glandular cells, endothelium, and mast cells. They effectively exert
competitive antagonism of histamine for H1-receptors. Itching and sneezing are
suppressed by antihistamine blockade of H1-receptors on nasal sensory nerves.
Therefore, antihistamine therapy represents a major therapeutic option (Monroe
et al., 1997). Antihistamines are used to treat allergies, motion sickness,
certain types of headaches, Crohn's disease, acute multiple sclerosis, and some
stomach secretory conditions. Antihistamines, which are well absorbed from the
gut and are metabolized primarily in the liver, act over a period of about 4-6
hours. They are available as OTC drugs or as prescription drugs. The following
are a list of common antihistamines available in the United States (Altruis,
2002):
Azatadine (Optimine®)
Carbinoxamine/Pseudoephedrine (Cardec®)
Cetirizine (Zyrtec®)
Cyproheptadine (Periactin®)
Dexchlorpheniramine (Polaramine®, Dexchlor ER®)
Fexofenadine (Allegra®)
Loratadine (Claritin®)
Promethazine (Phenergan®)
Tripelennamine (Pelamine®, Triplen®)
Histamine Deficiency
Histamine stimulates the production of digestive enzymes and gastric acid, which
aid in protein and fat digestion. A histamine deficiency can cause the body
to become more dependent on carbohydrates, which are used to synthesize cholesterol.
Therefore, increased carbohydrates and impaired fat metabolism can lead to high
total cholesterol levels. Histamine also stimulates the production of melatonin,
which is necessary for fat metabolism, in the pineal gland. A histamine deficiency
leads to lower levels of melatonin and decreased fat metabolism in the brain,
which can cause stress on the body. Whereas histamine is a stress regulator,
a deficiency in histamine leading to stress actually increases the body’s
need for histamine; this contributes to even greater worsening of symptoms (Prokarin,
2003). Studies have shown that histamine deficiency leads to poor folic acid
status (Kamsteeg, 2003). Interestingly, patients with multiple sclerosis are
deficient in histamine; in MS, the myelin sheath that insulates nerves of the
CNS and spinal cord is destroyed, and nerve fibers “short circuit”.
Histamine, which stimulates repair by increasing the production of myelin, is
greatly reduced in MS patients. Various histamine patches and replacement therapies
have been developed due to this information (Berger, 2000).
Type 1 Hypersensitivity: Immediate and Anaphylactic
Immediate hypersensitivity, commonly known as an allergic reaction, is the result
of an innocuous substance (ie. grass pollen, dead house-dust mites, certain
foods, or drugs) coming into contact with respiratory or gastrointestinal mucosa
(Rang, et al., 1995). Non-allergic individuals produce mainly TH1 cells and
low levels of IgG antibodies and respond weakly to these allergens. Atopic,
or allergic, individuals produce mostly TH2 cells and large quantities of IgE
antibodies that bind to receptors on mast cells (Spiegelberg, 1989). This overproduction
of IgE antibodies in response to otherwise innocuous antigens is responsible
for allergic reactions. The release of mediators, such as histamine, can cause
a number of effects localized to the nose (hay fever), bronchial tree (initial
phase of asthma), skin (urticaria), or gastrointestinal tract. A generalized
reaction to mediator release leads to anaphylactic shock (Rang et al., 1995).
Anaphylaxis was studied in the early 1900’s when a French physician, Charles
Richet, intended to test how much Portuguese Man of War toxic was required to
kill a dog. Although he was able to determine the lethal dose, many of the dogs,
which were not given the lethal dose, survived the experiments. Richet injected
the dogs a second time to determine whether they had become immune to the effects
of the first dose. Although he proposed that the first dose would provide protection
(prophylaxis) against the second injection, all of the dogs died. These opposite
effects were termed “anaphylaxis”. Later studies showed that anaphylaxis
is caused by the massive degranulation of mast cells and subsequent high levels
of histamine release into the tissues. Blood volume is reduced so severely that
the heart can no longer pump efficiently; at the same time, smooth muscle contraction
around the trachea can cause suffocation (Sompayrac, 1999).
References
Abbas, A., Lichtman, A., Pober, J. 1994. Cellular and Molecular Immunology.
Philadelphia, PA: W. B. Saunders Company. p. 287.
Altuis. Antihistamine.org. http://www.antihistamine.org. Accessed 3/10/2003.
Bach, Jean-Francois. 1982. Immunology. New York, NY: John Wiley & Sons,
Inc. p.
354-359, 459, 729, 745.
Berger, A. Tahoma Clinic. Histamine Patch.
http://www.tahoma-clinic.com/berger3.shtml. Accessed 3/10/2003.
Froese, A. 1980. Structure and function of the receptor for IgE. Critical Reviews
in
Immunology 1(2): 79-132.
Gallin, J., Weinstein, A., Cramer, E., and Kaplan, A. 1980. Histamine modulation
of
human eosinophil locomotion in vitro and in vivo. In: The Eosinophil in Health
and Disease, edited by A. A. F. Mahmond and K. F. Austen. New York, NY:
Grune and Stratton. p. 185-206.
Goldie, R. 1990. Receptors in asthmatic airways. The American Review of Respiratory
Disease 141(3): S151-6.
Kamsteeg, J., PhD. HPU-home. The association between histamine level, sun
sensitivity, and age in HPU-patients. http://www.hputest.nl.ehisres.htm. Accessed
3/10/2003.
Metcalfe, D., Baram, D., Mekori, Y. 1997. Mast cells. Physiological Reviews
77(4):
1033-79.
Monroe, E., MD, Daly, A., and Shalhoub, R., MD. 1997. Appraisal of the validity
of
histmine-induced wheal and flare to predict the clinical efficacy of antihistamines.
The Journal of Allergy and Clinical Immunology 99(2): S789-806.
Paul, William. 1984. Fundamental Immunology. New York, NY: Raven Press. p.
716-
720.
Prokarin. EDMS. The Official Prokarin Resource Site. Important new research
information for Prokarin Patients. http://www.edmsllc.com/diet.htm. Accessed
3/10/2003.
Rang, H., Dale, M., Ritter, J., Gardner, P. 1995. Pharmacology. New York, NY:
Churchill Livingstone. p. 226-229.
Sompayrac, Lauran, PhD. 1999. How the Immune System Works. Malden, MA:
Blackwell Science, Ltd. p. 37-38, 88.
Spiegelberg, H. 1989. Biological role of different antibody classes. The International
Archives of Allergy and Applied Immunology 90(1): 22-7.
email questions or comments
visit the official webpage of Davidson
College