*This website was produced as an assignment for an undergraduate course at Davidson College.*
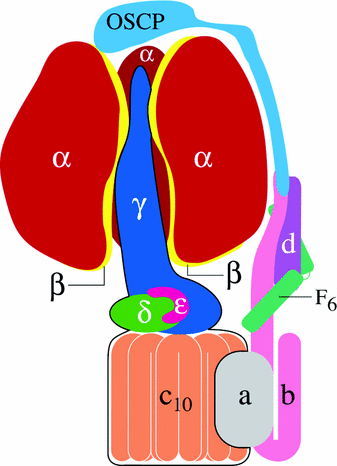
Yeast ATP Synthase
Mitochondrial ATP synthases in eukaryotes are more complex than the prokaryotic enzyme, especially with respect to the intramembrane F0 region. The yeast ATP synthase has 21 subunits, 14 of which serve to synthesize and hydrolyze ATP and seven of which are structural in nature. Yeast have four subunits in their F1 domains that correspond in form and function to the α, β, γ, and ε subunits in the E. coli enzyme, which are known as α, β, γ, and δ. The yeast rotor stalk contains an ε subunit completely different from the subunit of the same name in E. coli. Nine subunits in the F0 domain (4, OSCP, 6, d, 8, 9, f, h, and i/j) are necessary for ATP synthase function. When the gene for one of these chains is experimentally removed, the yeast cell’s respiration is inhibited. Of the above subunits, 4,6, and 9 resemble b, a, and c in E. coli (Ackerman and Tzagoloff, 2005).
From here forward, yeast subunits will be referred to by their E. coli equivalents for clarity. Yeast F0 proton channels consist of one a and ten c chains, while the other subunits form the stabilizing peripheral stalk. The δ chain in E. coli is replaced by OSCP in yeast, which stands for oligomycin-sensitivity conferring protein. This protein sits on top of the peripheral stalk touching the F1 domain. In bacteria, the δ peptide associates with the purified F1 domain, but in yeast it is firmly attached to the F0 region. While E. coli has two b subunits attached to its F0 domain, yeast mitochondria only have one, and d and h subunits add the stability that the second b subunit gives to the bacterial stalk. The yeast b subunit also differs from the bacterial protein in that it has two transmembrane regions instead of one. Subunits f and i/j form a hydrophobic foundation for the stator stalk and have their carboxyl termini in the intermembrane space and one transmembrane region each. Subunit 8 lies adjacent to the proton channel and the peripheral stalk and, thus, connects the proton channel to the motor element, the rotor stalk (Ackerman and Tzagoloff, 2005).
Subunits e, g, and k associate with the yeast ATP synthase only when it forms dimers. Since ATP synthase can function as a monomer or a dimer, these subunits are not necessary for cellular respiration. Dimers form when the coil-coil region of the e peptide interacts with subunits g and k. Transmembrane subunits e and g have hydrophilic regions in the intermembrane space, while the k subunit associates with them and is attached to the inner membrane (Ackerman and Tzagoloff, 2005).
Evolution of ATP Synthase
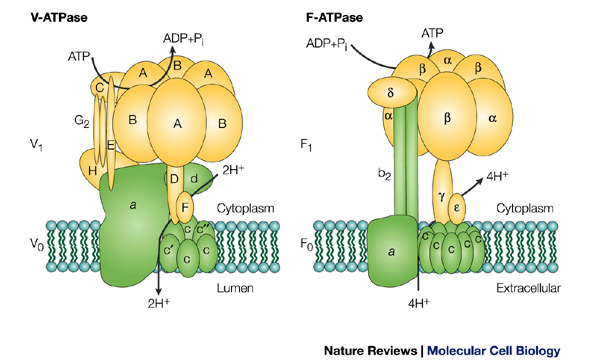
Three broad categories of ATP synthases have evolved: F-ATPases, A-ATPases, and V-ATPases. Most of the ATP synthases in eukaryotes and bacteria are F-ATPases, which have been discussed in detail on this website and consist of a cytoplasmic F1 domain and an intramembrane F0 domain. Archaea have a different type of ATP synthase, called A-ATPase. A-ATPases also contain two broad domains, designated A1 and A0, and have nine subunits. These nine subunits (designated A, B, C, D, E, F, H, I, and K) are not well characterized and the actual number and orientation of each is not known. Only the A and B subunits have been relatively thoroughly studied. There are three copies of each subunit oriented in an alternating, circular pattern in the “head” portion of A1, just as in the F-ATPases. Both F- and A-ATPases make ATP using the energy provided by an ion gradient and couple ATP synthesis to the movement of either H+ or Na+ ions down this concentration gradient through an intramembrane ion channel. V-ATPases are found in vacuoles and are different from the other two types of ATP synthases due to the fact that they only hydrolyze, not synthesize, ATP. That is, they break down ATP to ADP and Pi. The V-ATPases hydrolyzing ATP releases the energy necessary for the tasks that vacuoles perform, including endocytosis, bone reabsorption and acid hydrolase activation. V-ATPases, like the other two types of ATP synthases, have two domains, called V1 and V0. They have 13 subunits in the stoichiometry A3:B3:C:D:E:F:Gy:Hz:a:d:c:c’:c”. In addition to the fact that V-ATPases only hydrolyze ATP in vivo, they also differ from F- and A-ATPases in that the V1 and V0 can dissociate from each other, which produces decreased enzyme activity and, thus, reduced ATP hydrolysis. This reversible dissociation is clearly an enzyme control mechanism and has been observed in the laboratory during times of cellular stress, such as glucose starvation in yeast, when the cell needs ATP (Muller, Gruber, 2003).
All ATP synthase enzymes have the same basic structure composed of two major domains, each comprised of multiple subunits, joined by a rotor stalk and sometimes a peripheral stalk. This high degree of physical similarity suggests that all ATPases evolved from a single ancestor. Over the course of evolution, one gene was reproduced to create the multiple α and β subunits in F-ATPases and the A and B subunits in A- and V-ATPases. Mutations in this gene in the ancestors of the A- and V-ATPases removed catalytic capability from one of the B subunits in the “head” portion of the A1 and V1 domains. To fold correctly and maintain the overall function of the “head” region, however, this non-catalytic B peptide must still bind ADP or ATP. Such mutations also made the catalytic B subunits of A- and V-ATPases larger than the non-catalytic A subunits, unlike F-ATPases, in which the catalytic subunits are smaller than the non-catalytic ones (Muller, Gruber, 2003).
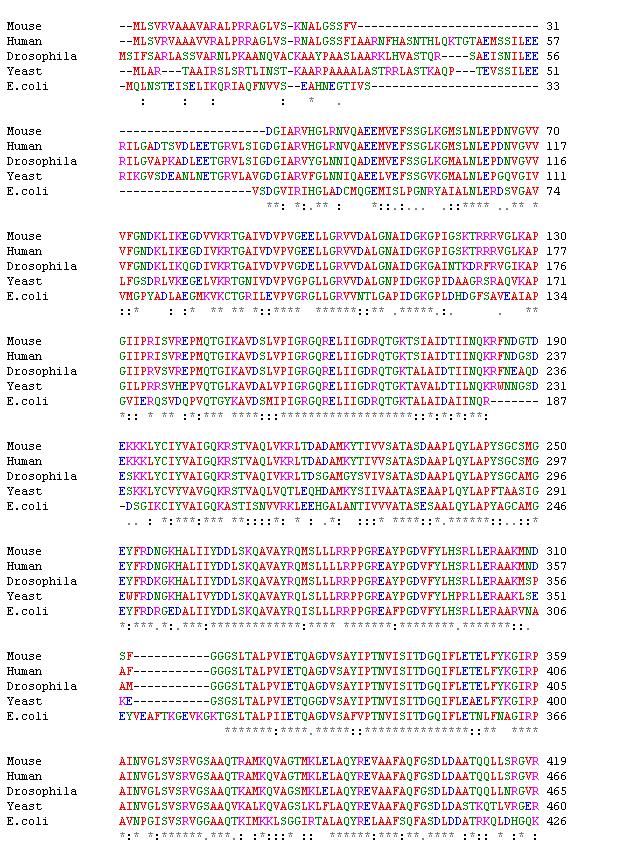

The α subunit of the F-ATPase is highly conserved, which is intuitive considering that the catalytic activity of the β subunits depends on contact with and, thus, proper folding of the α subunits. The comparison of the amino acid sequences from the F-ATPase of Escherichia coli bacteria, yeast (Saccharomyces cerevisiae), fruit flies (Drosophila melanogaster), mice(Mus musculus), and humans (Homo sapiens) shown in Figure 3 demonstrates that many of the amino acid residues are exactly the same across all five species. There are also many conservative substitutions, which do not dramatically alter protein folding, because they exchange one amino acid for another one in the same category. The mitochondrial ATP synthase α peptides in yeast, fruit flies, and humans are longer than E. coli, while the mouse α subunit has experienced some deletions and is shorter than that of E. coli (NCBI; ClustalW).

As mentioned above, all ATPases share a very distant common ancestor. On the phylogram in Figure 4, E. coli bacteria and yeast share a common ancestor, but the length of the lines from the branch point are rather long, which indicates that the ATP synthases in these two species have been evolving independently for a long time. This assumption makes sense given the fact that E. coli are prokaryotes, while yeast are eukaryotes. Mice and humans, on the other hand, are both mammals, so it makes sense that their ATPases share a very evolutionarily recent common ancestor. Fruit flies, the only insect being compared, understandably occupy a unique evolutionary branch from the other four species (NCBI; ClustalW).
Works Cited
Ackerman SH, Tzagoloff A. 2005. Function, structure, and biogenesis of mitochondrial ATP synthase. Progress in Nucleic Acid Research and Molecular Biology 80: 95-133.
Alpha subunit of the F1 sector of mitochondrial F1F0 ATP synthase, which is a large, evolutionarily conserved enzyme complex required for ATP synthesis; phosphorylated [Saccharomyces cerevisiae] [Internet]. Bethesda (MD): National Center for Biotechnology Information; 2009 Dec 9 [cited 2010 Mar 9]. Available from http://www.ncbi.nlm.nih.gov/protein/NP_009453.1?ordinalpos=1&itool=EntrezSystem2.PEntrez.Sequence.Sequence_ResultsPanel.Sequence_RVDocSum
ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit, isoform 1, isoform CRA_h [Mus musculus] [Internet]. Bethesda (MD): National Center for Biotechnology Information; 2007 Jun 7 [cited 2010 Mar 9]. Available from http://www.ncbi.nlm.nih.gov/protein/EDL09451.1?ordinalpos=2&itool=EntrezSystem2.PEntrez.Sequence.Sequence_ResultsPanel.Sequence_RVDocSum
ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit 1, cardiac muscle [Homo sapiens] [Internet]. Bethesda (MD): National Center for Biotechnology Information; 2007 Mar 9 [cited 2010 Mar 9]. Available from http://www.ncbi.nlm.nih.gov/protein/AAH39135.2?ordinalpos=5&itool=EntrezSystem2.PEntrez.Sequence.Sequence_ResultsPanel.Sequence_RVDocSum
ClustalW2 [Internet]. European Bioinformatics Institute; 2010 [cited 2010 Mar 9]. Available from http://www.ebi.ac.uk/Tools/clustalw2/index.html
F1 sector of membrane-bound ATP synthase, alpha subunit [Escherichia coli str. K-12 substr. MG1655] [Internet]. Bethesda (MD): National Center for Biotechnology Information; 2010 Mar 5 [cited 2010 Mar 9]. Available from http://www.ncbi.nlm.nih.gov/protein/NP_418190.1?ordinalpos=1&itool=EntrezSystem2.PEntrez.Sequence.Sequence_ResultsPanel.Sequence_RVDocSum
Gledhill JR, Walker JE. 2006. Inhibitors of the catalytic domain of mitochondrial ATP synthase. Biochemical Society Transactions 34: 989-992.
Mitochondrial ATP synthase alpha subunit precursor [Drosophila melanogaster] [Internet]. Bethesda (MD): National Center for Biotechnology Information; 2005 Apr 18 [cited 2010 Mar 9]. Available from http://www.ncbi.nlm.nih.gov/protein/CAA69202.1?ordinalpos=1&itool=EntrezSystem2.PEntrez.Sequence.Sequence_ResultsPanel.Sequence_RVDocSum
Muller V, Gruber G. 2003. ATP synthases: structure, function and evolution of unique energy converters. Cellular and Molecular Life Sciences 60: 474-494.
Nishi T, Forgac M. 2002. The vacuolar (H+)-ATPases--nature's most versatile proton pumps. Nature Reviews Molecular Cell Biology 3(2):94-103.