This web page was produced as an assignment for an undergraduate course at Davidson College. My Favorite Yeast Genes: Cox17 and the Non-annotated YLL007C |
Introduction: This web page investigates two yeast genes, Cox17, a previously annotated yeast gene, and YLL007C, a non-annotated yeast gene on the same chromosome. Both genes are located on the Crick strand of Chromosome XII in the S. Cerevisiae genome. The goal of this web page is to combine information from databases such as NCBI, NCBI Blast, Enterez, and PREDATOR to fully characterize the Cox17 yeast gene. In addition, this web page seeks to use those same databases to make inferences about the cellular role of the non-annotated YLL007C gene. Comprehensive information about Cox17 and YLL007C can be found at the Saccharomyces Genome Database. |
Characterization of Cox17 Cox17 Gene Ontology: Molecular Function: S. Cerevisiae Cox17 is a copper metallochaperone that transports copper from the cytosol to the mitochondrial inter-membrane space for delivery to cytochrom c oxidase. Cox17 transfers Cu to Sco1 as an intermediate step in Cu donation to cytochrome oxidase. Cytochrome c oxidase is the terminal complex of mitochondrial and bacterial respiratory chains. The cytochrome c oxidase utilizes the central iron of the Heme A prosthetic groups of cytochromes a and a3, each of which has an associated copper center. The synthesis of Heme A occurs in the inner mitochondrial membrane, however, copper must be imported from the cytoplasm. It is unlikely that copper ever exists as a free solute in yeast and it is known that external copper is imported into the cell via an ATP-dependent pump endoded by the CTR1 gene. It is believed that Cox17 is responsible for the delivery of this copper in the cytoplasm to the mitochondria (Glerum et al., 1996). Cox 17 transports this copper by binding as a binuclear cuprous-thiolate cluster.
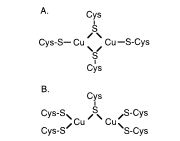
Figure 1: Candidate models for the binuclear Cu(I)-thiolate cluster in Cox17, (Permission pending from Srinivasan et al. 1998). Biological Process: Cox17 is involved in cytochrome c oxidase biogenesis and intra-cellular copper ion transport. Cox17 has been shown to be essential for the expression of cytochrome oxidase in Saccharomyces cerevisiae (Beers et al. 1997).
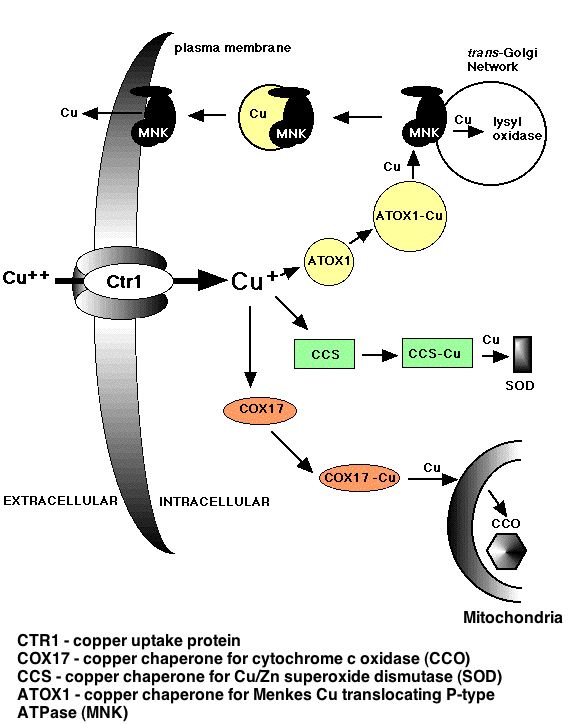
Figure 2: The mechanism of importation of copper ions from the extra-cellular space to cytochrome c oxidase in the mitochondria. Cox17 binds the copper ions once they have entered the cytoplasm. Permission pending from www.genetics.unimelb.edu.au/.../ projAfig.html Cellular Location: Cox17 is a soluble protein found in the cytoplasm and intermembrace space of the mitochondria in yeast. Other Useful Gene Information: Comprehensive gene information about Cox17 can be found at Enterez Gene. Gene ID for Cox17: 850651 Nucleotide Sequence: ATGACTGAAACTGACAAGAAACAAGAACAAGAAAACCACGCGGAGTGCGAGGACAAACCT Complete Chromosome XII Sequence: NC_001144 Cox17 Chromosomal Location: Cox17 is located on the Crick strand of chromosome XII in S. Cerevisiae.
Figure 3: Chromosomal location of Cox17 on the Crick strand of Chromosome XII. Cox17 is flanked by two neighboring genes: PSR1, a plasma membrane Sodium Response 1 protein, and YLLOO7C, an non-annotated yeast gene. Figure obtained from Saccharomyces Genome Database. Blastn Search: A Blastn search was performed using the Cox17 genomic DNA sequence shown above. The results are seen below:
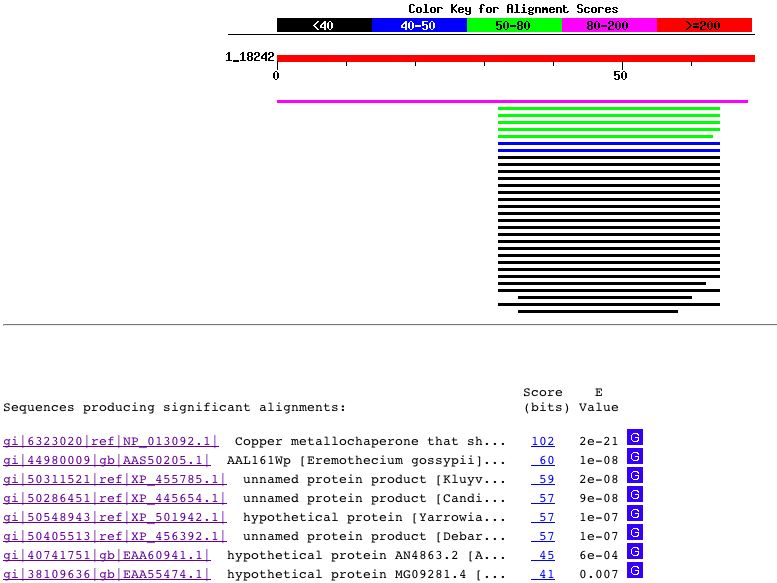
Figure 4: Blastn results for the genomic DNA sequence of Cox17. Figure obtained using the Blastn database. The only sequences producing significant alignments with Cox17 genomic DNA were those within the yeast genome that actually contained the Cox17 sequence. For example, one of the hits was the genomic sequence for all of Chromosome XII. Protein Information: Cox17 is a low molecular weight protein found in both yeast an mammalian cells (Beers et al., 1997). The Cox17 ORF in yeast encodes an 8057 Dalton polypeptide (Srinivasan et al. 1998). The yeast Cox17 contains seven cysteinal residues, six of which are conserved in the human Cox17 protein. Cox17 Amino Acid Sequence: MTETDKKQEQENHAECEDKPKPCCVCKPEKEERDTCILFNGQDSEKCKEFIEKYKECMKG NCBI Protein Accession Number: NP_013092 Blastp Searches: Using the Cox17 amino acid sequence shown above, a search of the Blastp database yielded the following results:
Figure 5: Blastp results for the Cox17 protein sequence. Figure was obtained using the Blastp database. Most of the significant alignments between the S. Cerevisiae Cox17 protein and other proteins are either unnamed or hypotheitcal protein products of various fungal species. The Blastp results also indicated that the Cox17 protein contained only one conserved domain:
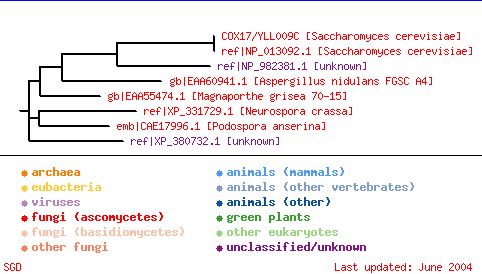
Figure 6: NCBI conserved domain search for the Cox17 protein. Figured obtained from NCBI Conserved Domain. The Cox17 domain is conserved in 24 eukaryotic species and is known to be involved in copper transport from the cytosol to the inter-membrane space of the mitochondria in all of these speices. In order to determine if there were any more broadly related proteins, a PSI-Blast analysis of the Cox17 yeast protein was performed. PSI-Blast yields the following dendogram:
Figure 7: Dedogram generated using PSI Blast analysis of the Cox17 protein. PSI blast identifies broadly related protein families. This figure was obtained from the Saccharomyce Genome Database under the information for Cox17. This indicates that Cox17 is most closely related to other yeast and fungal proteins, showing some similarity with other unclassified/unknown sequences. Mutations in Cox17: Yeast cells that harbor a mutant Cox17 gene are respiratory-deficient but are still able to synthesize mitochondially and nuclearly encoded cytochrome oxidase units. This respiratory defect can be partially rescued by high exogenous Cu(II) levels, providing further evidence that Cox17 in involved with copper ion delivery to the mitochondria. A study of the effects of various mutations in the Cox17 gene revealed that three of the seven cysteinyl residues are critical for Cox17 function, and these three residues are present in a Cys-Cys-Xaa-Cys sequence motif. Single substitution of any of the three cysteines with serines results in a nonfunctional cytochrome oxidase complex. Mutant proteins that lack only one of the critical Cys residues retain the ability to bind three Cu(I) ions and are imported within the mitochondria. In contrast, Cox17 molecules with a double Cys-Ser mutation exhibit no Cu(I) binding but are still localized to the mitochondria (Heaton et al. 2000). Homologs of Cox17 There are several known homologs of yeast Cox17 in various other organisms. Some of their protein accession numbers are listed below: Human: NP_005685 Mouse: P56394 Rat: NP_445992 Arabidopsis thaliana: NP_566508 Cox17 in Mammalian Cells: A study was completed by Amaravadi et al. in 1997 to determine if Cox17 is present in mammalian cells. To make this determination, the group used a yeast straicontaining a null mutation in Cox17 and transformed this with a human cDNA expresion library. All of the clones obtained from the transformations contained a common cDNA sequence homologous to yeast Cox17. This cloning of the mammalian Cox17 homolog suggests that this protein plays a role in copper recruitment in alleukaryotic cells. This gene is also a target being investigated in cytochrome oxidase definciencies in humans. Cox23: Another yeast protein, Cox23, has been shown to be required for the expression of cytochrome oxidase. Cox 23 is homologous to Cox17 and null mutants can be rescued by high exogenous copper concentrations when the mutant contains Cox17 on a high copy plasmid. Like Cox17, Cox23 is found in the intermembrane space of the mitochondria, suggesting that they function in a common pathway (Barros et al. 2004). |



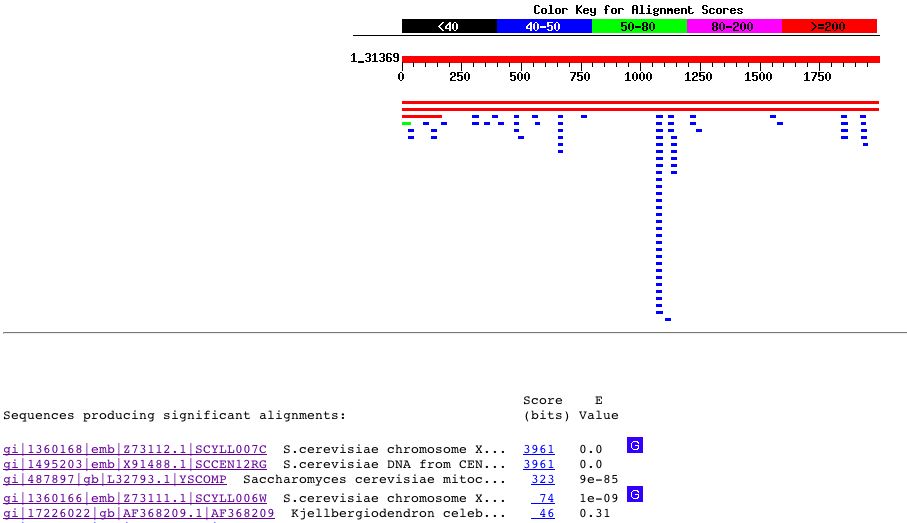
Characterization of YLL007C YLL007C is considered to be a hypothetical ORF on chromosome XII of S. Cerevisiae. YLL007C Gene Ontology: Molecular Function: According to information provided by the Saccharomyces Genome Database, YLL007C has no known molecular function. Cellular Process: Similarly, the cellular processes of the YLL007C gene are also unknown. Cellular Location: According to the SGD, YLL007C is a cytoplasmic protein.Other Useful Gene Information: Comprehensive Gene Information about YLL007C can be found at Enterez Gene Genomic DNA Sequence of YLL007C : ATGAAGCATAACAGGCAGATGCCAGATGAAAGCTTGGAAAACATAAAAGTGCTACTGAAT Chromosomal Location: YLL007C is located on the Crick strand of Chromosome XII. See Figure 3 above. Blastn Search: Using the genomic DNA sequence for YLL007C listed above, I performed a Blastn search. The results can be seen below:
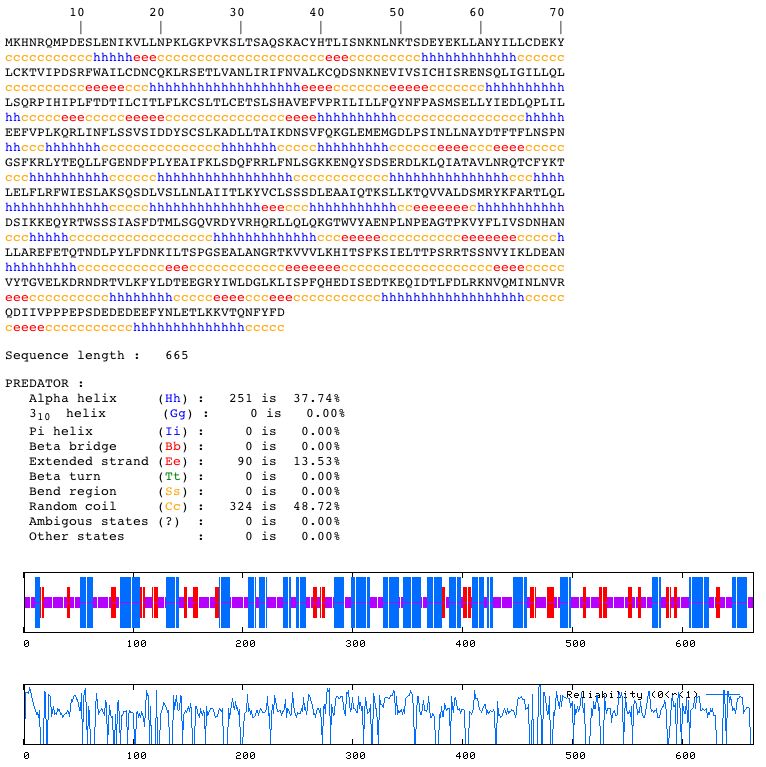
Figure 8 : Blastn results for the genomic DNA sequence of YLL007C. This figure was obtained using the Blastn database. Just as with the sequence for the Cox17 gene, the only significant matches were those within the yeast genome. Once again, these hits contained overlapping regions of S. Cerevisiae Chromosome XII. Protein Information: YLL007C Amino Acid Sequence: 1 mkhnrqmpde slenikvlln pklgkpvksl tsaqskacyh tlisnknlnk tsdeyeklla Protein Accession Number: NP_013094 Blastp Searches: With the amino acid sequence for YLL007C listed above, I performed a Blastp search:
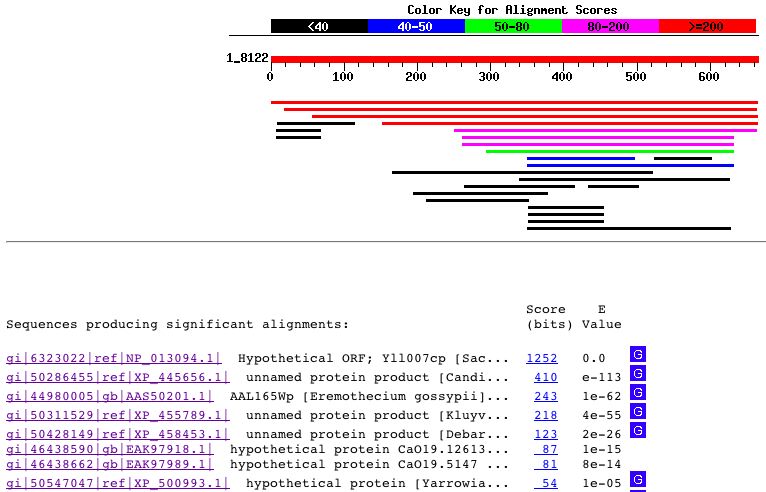
Figure 9 : Blastp results for YLL007C amino acid sequence. This figure was obtained using the Blastp database. The only significant protein similarities that I found were unnamed or hypothetical proteins that had no known functions. With little information provdided from the Blastp results, I decided to perform a conserved domain search for this protein. The results of this conserved domain search can be seen below:
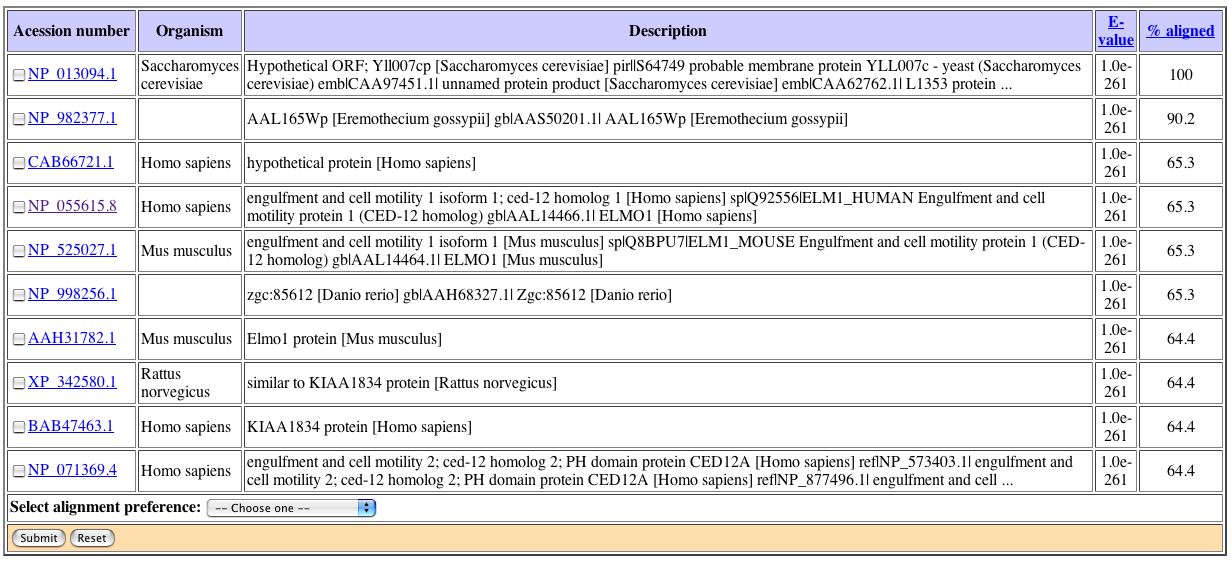
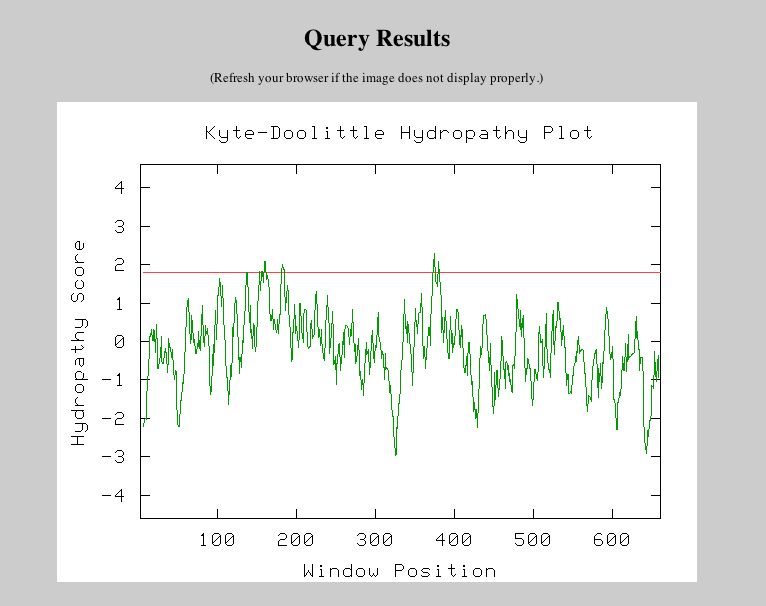
Figure 10: Results of the Conserved Domain search using the amino acid sequence from YLL007C. This figured was obtained from NCBI's Conserved Domain Database. Unfortunately, there were no hits found. Next, I decided to investigate the PSI Blast data for YLL007C on the Saccharomyces Genome Database website. PSI Blast Results: |

|
{kind=link}