
Effects of Pollen Addition on Andromonoecy in Horsenettle (Solanum carolinense )
Janet C. Steven and Patricia A. Peroni
Department of Biology, Davidson College
Abstract
We examined the effects of pollen addition
and flower number on fruit and seed set in Solanum carolinense (Solanaceae),
a weedy perennial commonly known as horsenettle. Individual plants bear
both male and hermaphroditic flowers, a breeding system referred to as andromonoecy.
The questions investigated include:
1. Are fruit and seed production in horsenettle populations pollen limited?
2. Do pollen additions also affect the production of male and hermaphroditic
flowers?
3. Is total flower number or the percentage of flowers that are male on
a plant related to percent fruit set, and therefore possibly related to
the attraction of pollinators?
We conducted pollen additions on 25 plants in each of two populations, and
also monitored 25 control plants at each site. The mean percent fruit set
and seed set per fruit of the pollen addition treatment was significantly
greater than that of the control, which indicates that both populations
are pollen limited. Fruit production did not correlate with total flower
number or percent male flowers, which implies that a larger floral display
does not affect the receipt of compatible pollen from pollinators. These
results suggest that male flowers may provide increased opportunities for
plants to sire offspring, either by allowing more cost effective pollen
production or more efficient uptake of pollen by bees than hermaphroditic
flowers. The percent of flowers on a plant that were male was significantly
greater in the pollen addition treatment than in the control, which suggests
that pollen addition affects gender expression in horsenettle. Pollen limitation
may select for an increase in pollen or flower production, and andromonoecy
may provide the flexibility in resource allocation that makes such an increase
possible.
METHODS
* Sampled two populations in Giles Co,
VA - Givens and Domermouth
* Randomly assigned plants to treatment:
-pollen addition
-open pollinated control
* Monitored study plants daily for 3 wk and, then every other day for
2 wk
-marked all flowers as opened and recorded sex
-applied pollen to all open hermaphroditic flowers on plants assigned to
pollen addition treatment
-recorded eventual fate of each flower
-measured height of each plant and density of conspecifics
* Collected mature fruits from study plants at end of growing season
-dried to constant mass and weighed
-counted seed number for each of 20 randomly selected fruits
Hermaphroditic horsenettle flower. Note the large functional pistil that extends beyond the anthers.

Male horsenettle flower. Note the
absence of a functional pistil.
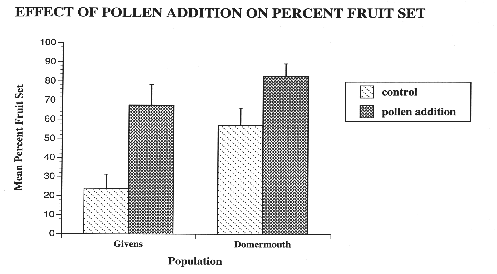
Figure 1: Mean fruit set of plants on finished inflorescences in pollen addition and control treatments in two horsenettle populations. Two-way ANOVA results indicate significant population (F=27.50; df=1,88; p=0.0001) and treatment (F=45.47; df=1,88; p=0.0001) effects. Fruit set is expressed as the percentage of hermaphroditic flowers that set fruit on inflorescences that finished flowering. In the pollen addition treatment, n=23 and n=25 for Domermouth and Givens, respectively. For the control, n=26 in both populations. Error bars represent 2 standard error from the mean.
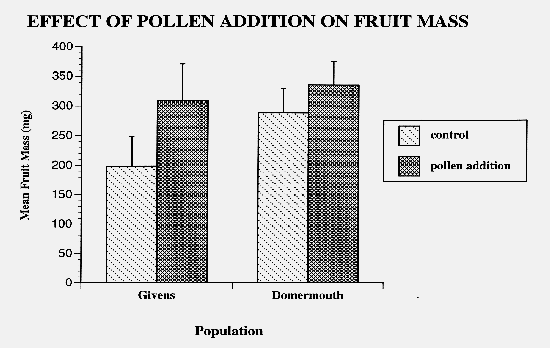
Figure 2: Mean of the average fruit mass per plant in mg in two horsenettle populations. Two-way ANOVA results indicated significant population (F=5.79; df=1,79; p=0.0185) and treatment effects (F=10.85; df=1,79; p=0.0015). All fruits were set by flowers open during the manipulations. Mature fruits were collected and dried to constant mass. In the pollen addition treatment, sample size was n=18 and n=20 for Givens and Domermouth, respectively. In the control treatment, sample size was n=20 and n=22 in the Givens and Domermouth populations respectively. Error bars represent 2 standard error from the mean.
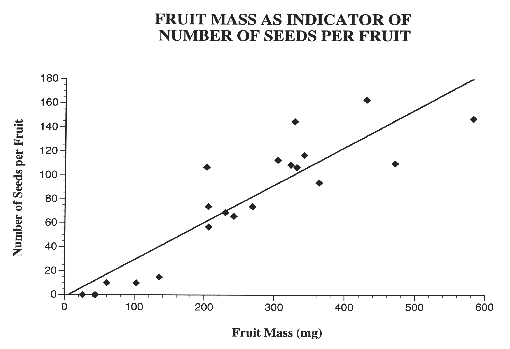
Figure 3: Fruit mass vs. number of seeds per fruit. Number of seeds per fruit showed a significant regression with fruit mass (Rsquare= 0.7996, F=471.84, df=1,18, p<0.0001). Best fit line: number of seeds= -0.7698 + 0.312089*(fruit mass). N=20 fruits.
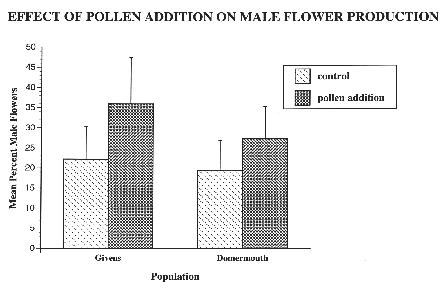
Figure 4: Mean percent male flowers on finished inflorescences in pollen addition and control treatments in two horsenettle populations. Results of an ANCOVA that used plant height and density of conspecifics as covariates indicate a significant treatment effect (F=5.69; df=1,86; p=0.0192). In the pollen addition treatment, n=23 and n=25 for Domermouth and Givens respectively. For the control, n=26 in both populations. Error bars represent 2 standard error from the mean.

Figure 5: Percent fruit set vs. total flower number and percent male flowers for plants in control treatment only. Percent fruit set expressed as fraction of hermaphroditic flowers that set fruit. Percent male flowers represents the percentage of all flowers on a plant that were male. N=26 plants per population. Results of both Spearman and Pearson correlation analyses indicate no significant correlation between percent fruit set and either total flower number or percent male flowers.
CONCLUSIONS
* Both populations are pollen limited.
* Pollen addition was associated with greater seed set per fruit (since
fruit mass correlated significantly with seed number).
* Pollen addition was associated with a shift in gender expression.
* Fruit set was not associated with total flower number or the proportion
of male to all flowers.
* These results suggest that the potential role of male flowers involves
pollen donation, not pollinator attraction.
Male flowers may benefit plants if they:
- permit better or more efficient pollen uptake and transfer than bisexual
flowers
- provide a cost effective mechanism for increasing pollen supplies.
![]()
![]()
© Copyright 2000 Department of Biology, Davidson
College, Davidson, NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu